
Recherche:Métabolisme prébiotique

Premières réflexions après Origins2011
[modifier | modifier le wikicode]écrites le 20/07/11
Les processus physico-chimiques du liposome
[modifier | modifier le wikicode]- L'information agit sur un processus physico-chimique ce qui organise la structure (exemple: réaction du gradient protonique à un changement ionique du milieu externe, les champs électriques en déplacement réorganisent les petites molécules à la surface et dans la membrane).
- La nouvelle organisation renforce le processus ou en crée un nouveau (exemple: les zwittérions favorisent la formation et la pénétration des ionophores dans la membranes).
Processus quantiques
[modifier | modifier le wikicode]Les processus quantiques peuvent être à l'origine de l'excès de chiralité qui peut à son tour se propager par résonance. Le liposome par sa structure inter-réagit quantiquement avec l'environnement ce qui peut représenter la connaissance du soi, de là la théorie de la cognition pour l'initialisation du vivant.
Les mouvements de la molécule de phospholipide dans la membrane
[modifier | modifier le wikicode]- Mouvement latéral à la queue leu-leu ----> très favorable pour le rapprochement des aa attirés par les zwitterions.
- Mouvement rotationel ---> Chiralité du phosphate: changements rapides de l'état chiral à l'état achiral favorisant la violation de parité.
- Vibrations et torsions des queues hydrophobes ---> s'ajoute à l'encombrement stérique de la tête hydrophile pour la chiralité du glycérol. Ces mouvements doivent être très importants puisqe les queues sont longues.
- Mouvement de flexion mentionné dans wikipédia: à confirmer.
- Le processus de flip-flop: utilisé par le vivant pour faire passer les PLDs du feuillet interne au feuillet externe avec une flipase. Qu'en est-il sans flipase quand il y a hydrolyse d'une liaison ester de la tête hydrophile ou addition de PLD dans le feuillet externe, au-delà du flip-flop de base.
La formation des feuillets dans les vésicules aqueuses dans l'huile
[modifier | modifier le wikicode]Les vésicules aqueuses dans l'huile sont soumises à des processus physico-chimiques spécifiques qui leur sont propres: isolement et concentration du milieu réactionnel; les durées des réactions peuvent être infinies pour le feuillet interne et très longues pour le feuillet externe; la géométrie concave doit jouer sur l'encombrement stérique et la formation des têtes hydrophiles, donc sur la chiralité du glycérol; la phase huile peut influer sur l'ordonnancement des queues hydrophobes.
La chiralisation de la molécule de phospholipide lors de la formation du liposome
[modifier | modifier le wikicode]La chiralité du glycérol est soumise à 4 contraintes:
- l'encombrement stérique de la tête hydrophile;
- les forces de torsion et de vibration des queues hydrophobes longues; (J. Reisse)
- la résonance quantique du liposome entier;
- la double chiralité temporaire du phosphate.
La chiralité du phosphate varie de façon continue et répond rapidement aux sollicitations du milieu externe. La PVED du phosphate devrait dans ces conditions favoriser une des 2 formes énantiomériques par rapport à l'autre.
Le gradient protonique à travers la membrane
[modifier | modifier le wikicode]voir chimio-osmose prébiotique pour la formation de ces gradients par les ionophores, et l'anti-gradient imposé par la diffusion des H+ (nouvel article).
Le potentiel électrique à travers la membrane
[modifier | modifier le wikicode]voir chimio-osmose prébiotique. Le gradient protonique crée un potentiel électrique qui déplacera les électrons délocalisés de certaines molécules poly-insaturées comme les hydroquinones et fera évoluer les hèmes et d'autres coenzymes en chaîne d'oxydo-réduction.
Insertion des molécules hydrophobes dans la membrane
[modifier | modifier le wikicode]Ionophores, hèmes, quinones et noyaux aromatiques.
Diffusion des petites molécules à travers la membrane
[modifier | modifier le wikicode]- Bases nucléiques: diffusion facile car hydrophobes; une fois entrées le chemin vers l'ADN est plus facile que celui de l'ARN car la synthèse du dRibose est auto-catalytique.
- H2S: diffusion facile; nécessaire pour les coenzymes et surtous les liaisons thiols riches en énergie peuvent intervenir dans l'initialisation du métabolisme avant l'ATP.
- CH4: important pour la méthanotrophie pour la boucle métabolique des phospholipides qui peut s'établir indépendamment de la multiplication global du protobionte.
- aa hydrophobes (10) à l'origine du code génétique.
- Les acides carboxyliques petits et faiblement acides, donc peu polarisés pouvant traverser la membrane. Une fois à l'intérieur ionisation pour maintenir l'équilibre ionique avec l'extérieur, et réactions chimiques spécifiques de l'intérieur.
La molécule de phospholipide est un zwittérion
[modifier | modifier le wikicode]À développer: pKa du phosphate 0.8(PE) et 0.5(PC); pKa du NH2 9.6(PE), 9.8(PC).
Le liposome est un catalyseur
[modifier | modifier le wikicode]Catalyse du D-Ribose par L-Val2: S. Pizzarello (2009)[1]
Oligomérisation des aa par le liposome: H. Tsukahara (2001)[2]
L'homochiralité
[modifier | modifier le wikicode]Chez les bactéries et les archées l'homochiralité est séparée de l'extérieur ( par rapport à l'autre forme chirale) pour éviter la destruction de son effet quantique par une paroi racémique, structurée et rigide (ne rentrant pas en contact avec les protéines membranaires) ne laissant pas entrer les molécules de l'autre forme chirale.
Le RNA est l'intermédiaire entre le DNA et les protéines
[modifier | modifier le wikicode]Le RNA ne peut être libre
[modifier | modifier le wikicode]Il n'y a pas de bactérie ou d'archée à ARN (sans ADN). Les virus à ARN sont encapsulés dans une coque de protéines. Il n'y a que les tRNA qui sont libres encore sont ils chapeautés par un acide aminé. Les ribozymes appartiennent aux eucaryotes. Dans une bactérie ils désorganiseraient le métabolisme et toutes les expériences du RNA world concernent soit la réplication, la transcription et la manipulation des brins d'ADN ou d'ARN mais jamais la catalyse de réactions métaboliques. Il n'y a que le ribosome qui fait fonction de ribozyme pour la synthèse des protéines.
Transcription réverse, réplication, transcription puis traduction primitive
[modifier | modifier le wikicode]Si l'ARN libre (l'ARNm, sauf les tRNA) est rapidement dégradé, l'ADN simple brin ne se trouve jamais seul. Il est
- soit temporairement avec l'ARN lors de la transcription directe et réverse;
- soit avec les protéines pour l'initialisation de la transcription (ouverture locale du double brin);
- soit avec de petits simples brins d'ADN pour la réplication dans la fourche de réplication.
Par contre à l'inverse du RNA, l'ADN double brin est stable et peut exister seul sans protéines.
Il faut vérifier à ce sujet si la réplication des ribozymes (principe du réplicateur) sans l'aide de l'ADN ne pose pas un problème à la théorie du RNA world, en vue de ce qui se passe réellement chez le vivant.
Cette remarque que le RNA ne puisse être libre et qu’il ait besoin du DNA pour se répliquer met à plat la théorie du RNA world, mais aussi mon ébauche de la reproduction des protéines après réplication du RNA, dans la théorie de la chimio-osmose prébiotique. Or dans cette théorie la chélation du RNA par les protéines membranaires et les aa, est en continuité avec la formation de protéines fonctionnelles et constitue la mémorisation de ces fonctionnalités dans un autre support qui peut se répliquer. Si le RNA a besoin du DNA pour se répliquer, alors il faut supposer que ce dernier apparaisse en même temps et même avant le RNA.
On voit ainsi que si on privilégie la mémorisation d'une fonction peptidique dans le RNA, comme dans la théorie de la chimio-osmose prébiotique, puisque le DNA n'existe pas en simple brin pour la chélation, pour que ce dernier se réplique, il faut d’abord qu’il soit reverse-transcrit en DNA, que ce dernier se réplique puis transcrit en RNA. Le nouveau RNA peut être alors chélaté par des aa pour former à peu près la même protéine de départ, chélation qui évoluera en traduction.
J'utilise ci-dessus des concepts du vivant actuel, mais si on se met dans les premières étapes de ce processus des AN, ça sera d’abord des monomères puis des oligomères de RNA, DNA et peptides coopérant ensemble. Un système nettement élémentaire mais très évolutif. Il démarre en étant attaché à la membrane par les premières protéines membranaires, mais les fonctions peptidiques qui se créent sont différentes des protéines membranaires, car le processus central qui les sous-tendent est la chélation par liaison hydrogène aux AN.
Les investigations futures vont consister à rechercher les étapes logiques, c'est-à-dire en continuité avec la chimio-osmose prébiotique, qui feront évoluer le système des AN vers la complexité du vivant actuel. Or la théorie de la chimio-osmose prébiotique débouche sur l'initialisation du métabolisme, par la synthèse des sucres phosphates. Pour avoir des AN, dans la chimio-osmose prébiotique, les bases nucléiques hydrophobes sont produites à l'extérieur dans la poche de pétrole prébiotique, et rentrent dans le liposome par diffusion.
Dans des études antérieures sur la prébiotique, j’avais analysé le métabolisme du vivant actuel, pour essayer de chercher les réactions les plus simples susceptibles de l'initialiser. Le résultat de ces recherches c’est que le dRibose-5P est obtenu avec une seule réaction d'un seul enzyme et sans coenzyme, alors que le Ribose-5P nécessite 5 réactions, 4 enzymes dont un nécessitant du Zn++ et un de la vitamine B1 comme coenzyme.
Il apparaît dès lors qu’il est tout à fait plausible
- que les oligo DNA soient apparus avant les oligo RNA, ou en même temps,
- que les fonctions peptidiques du métabolisme nucléique naissant, soient peu différenciées pour distinguer dRibose et Ribose, et remplir, imparfaitement certes, les 3 fonctions de transcription directe et réverse et la réplication du DNA en même temps.
La pression d'évolution se faisant avec des olgo AN de plus en plus longs. Cette incapacité des distinguer dRibose du Ribose se trouve par exemple dans le fait que certains enzymes actuels peuvent utiliser du dATP à la place de l'ATP, bien que moins efficacement.
Sélection
[modifier | modifier le wikicode]On ne devrait jamais employer le mot sélection pour des molécules, car il entaché d'anthropomorphisme et concerne surtout la sélection darwinienne basée sur le tirage au hasard de séquences d'ADN caché par un phénotype. Il faudrait parler de regroupement par le processus quantique de Bose-Einstein ou par attirance par les forces faibles de Van der Wals, London et hydrogène. Un être vivant est unique, identifiable et mortel, alors qu'une molécule donnée n'a pas ces attributs et se recrée reversiblement par réaction chimique.
Premières étapes du métabolisme des acides nucléiques, AN
[modifier | modifier le wikicode]Les 1eres étapes du métabolisme
[modifier | modifier le wikicode]écrites le 22/07/11
Le principe c’est la recherche de processus évolutifs en progressant des processus physiques (quantique, électromagnétique, aux processus physico-chimiques (osmose, liaison ionique) puis chimiques (liaison H, ester et autres liaisons covalentes). En recherchant les 1eres enzymes j’espère débusquer ces processus (la fonction crée l'organe?), par exemple pour dATP on n'a pas besoin que l'adénine rentre à l'intérieur, l'enzyme peut être dans la membrane.
- les 1eres étapes:
- hydrolyse de la liaison ester: par hydrolase dans la membrane qui a dû être formée par le processus de chimio-osmose.
- de même pour la flipase, d'où accélération du flip-flop ( canaux Phosphate à développer).
- le métabolisme actuel montre qu'on obtient P-ethanolamine et P-glycerol mais pas de P-serine.
- la serine serait formée par carboxylation de l'éthanolamine à l'intérieur du liposome d'où l'origine de la longueur des aa.
- faire le tableau de la diffusion des bases nucléiques, de l'acétaldéhyde (éthanol), les 2 morceaux de B1, pyridoxine, nicotinamide et FMN (et biotine?).
- dRibose-5P, autocatalyse.
- Ribose-5P, autocatalyse: développer B1 et les 2 voies métaboliques des AN.
- réaction dA, dG, ... donne le P libre qui servira aux redox (chimio-osmose) pour former PP précurseur d'ATP.
- dA + PP donne dAMP puis dADP puis dATP qui remplace ATP.
- hydrolyse de la liaison ester: par hydrolase dans la membrane qui a dû être formée par le processus de chimio-osmose.
- Les processus: qu'est-ce qui pousse à l'hydrolyse de la liaison ester, au flip-flop, à la carboxylation de l'éthanolamine?
- L'hydrolyse(H2O) de la liaison ester P-ser donnerait de la ser libre, de même sa thiolyse(H2S) donnerait de la cys libre. On peut imaginer des radicaux (R-H) libérer le P. Il faut alors étudier la diffusion des aa dans la membrane ainsi que ces radicaux.
Les enzymes primitives
[modifier | modifier le wikicode]Écrit le 26/07/11
Recherche des enzymes dont je soupçonne qu’elles soient issues d'enzymes primitives, donc qui auraient initialiser le métabolisme (KEGG ): Synthèse des aa à l'intérieur du liposome, autres que les aa hydrophobes qui auraient participé aux ionophores.
Tryptophane:
[modifier | modifier le wikicode]On ne le trouve pas dans les synthèses abiotiques dans les conditions hydrothermales.
- EC 4.2.1.20 Cofactor: Pyridoxal phosphate. Indole ( serait hydrophobe ) Biosynthèse du Tryptophane
A pyridoxal-phosphate protein. The alpha-subunit catalyses the conversion of 1-C-(indol-3-yl)glycerol 3-phosphate to indole and glyceraldehyde 3-phosphate. The indole then migrates to the beta-subunit where, with serine in the presence of pyridoxal 5'-phosphate, it is converted into tryptophan. Also catalyses the conversion of serine and indole into tryptophan and water, and of 1-C-(indol-3-yl)glycerol 3-phosphate into indole and glyceraldehyde phosphate (the latter reaction was listed formerly as EC 4.1.2.8). In some organisms, this enzyme is part of a multifunctional protein, together with one or more other components of the system for the biosynthesis of tryptophan [EC 2.4.2.18 (anthranilate phosphoribosyltransferase ), EC 4.1.1.48 (indole-3-glycerol-phosphate synthase), EC 4.1.3.27 (anthranilate synthase) and EC 5.3.1.24 (phosphoribosylanthranilate isomerase).
L-Serine + Indole => L-Tryptophan + H2O
L-Tryptophan + D-Glyceraldehyde 3_P + H2O <=> L-Serine + Indoleglycerol_P
Indoleglycerol_P <=> Indole + D-Glyceraldehyde 3_P
- EC 4.1.1.48 Cofactor: Pyruvate
In some organisms, this enzyme is part of a multifunctional protein, together with one or more other components of the system for the biosynthesis of tryptophan [EC 2.4.2.18 (anthranilate phosphoribosyltransferase), EC 4.1.3.27 (anthranilate synthase), EC 4.2.1.20 (tryptophan synthase) and EC 5.3.1.24 (phosphoribosylanthranilate isomerase).
Indoleglycerol_P +
CO2
+ H2O <=> 1-(2-Carboxyphenylamino)-1-deoxy-D-ribulose 5_P
- EC 5.3.1.34 Cofactor: néant
In some organisms, this enzyme is part of a multifunctional protein, together with one or more other components of the system for the biosynthesis of tryptophan [EC 2.4.2.18 (anthranilate phosphoribosyltransferase), EC 4.1.1.48 (indole-3-glycerol-phosphate synthase), EC 4.1.3.27 (anthranilate synthase) and EC 4.2.1.20 (tryptophan synthase).
1-(2-Carboxyphenylamino)-1-deoxy-D-ribulose 5_P <=> N-(5-Phospho-D-ribosyl)anthranilate
- EC 2.4.2.18 Cofactor: néant
In some organisms, this enzyme is part of a multifunctional protein together with one or more other components of the system for biosynthesis of tryptophan [EC 4.1.1.48 (indole-3-glycerol-phosphate synthase), EC 4.1.3.27 (anthranilate synthase), EC 4.2.1.20 (tryptophan synthase) and EC 5.3.1.24 (phosphoribosylanthranilate isomerase).
N-(5-Phospho-D-ribosyl)anthranilate + PP <=> Anthranilate + 5-Phospho-alpha-D-ribose 1_PP (PRPP)
- EC 1.13.11.23 Cofacteur : Néant Dihydroxyindole ( serait hydrophobe ) Métabolisme du Tryptophane
2,3-Dihydroxyindole + Oxygen => Anthranilate +
CO2
- EC 1.14.16.3 Cofacteur : Fe++ Tetrahydrobiopterin ( serait hydrophobe )
Tetrahydrobiopterin + Anthranilate + Oxygen => 3-Hydroxyanthranilate + Dihydrobiopterin + H2O
- EC 1.13.11.6 Cofacteur : Fe++
3-Hydroxyanthranilate + Oxygen => 2-Amino-3-carboxymuconate semialdehyde
2-Amino-3-carboxymuconate semialdehyde => quinolinate réaction sans enzyme, en.wiki
- EC 2.4.2.19 Métabolisme du NAD
Quinolinate + 5-Phospho-alpha-D-ribose 1-PP => Nicotinate D-ribonucleotide + PP +
CO2
This is the first enzyme that prokaryotes and eukaryotes have in common in the production of NAD+ as some prokaryotes use an L-aspartate pathway to produce quinolinate whereas all eukaryotes use tryptophan as the starting material.
- EC 1.4.3.16 Cofacteur : FAD
L-Aspartate + Oxygen => Iminoaspartate + Hydrogen peroxide
A flavoprotein (FAD). L-Aspartate oxidase catalyses the first step in the de novo biosynthesis of NAD+ in some bacteria. O2 can be replaced by fumarate as electron acceptor, yielding succinate [5]. The ability of the enzyme to use both O2 and fumarate in cofactor reoxidation enables it to function under both aerobic and anaerobic conditions [5]. Iminosuccinate can either be hydrolysed to form oxaloacetate and NH3 or can be used by EC 2.5.1.72, quinolinate synthase, in the production of quinolinate. The enzyme is a member of the succinate dehydrogenase/fumarate-reductase family of enzymes.
- EC 1.4.1.21 Cofacteur : néant
L-Aspartate + NAD+ <=> Iminoaspartate + NADH + H+
- EC 2.5.7.72 Cofacteur : [4Fe-4S]
Iminoaspartate + Glycerone_P => Quinolinate + 2 H2O + P
An iron-sulfur protein that requires a [4Fe-4S] cluster for activity [1]. Quinolinate synthase catalyses the second step in the de novo biosynthesis of NAD+ from aspartate in some bacteria, with EC 1.4.3.16 (L-aspartate oxidase) catalysing the first step and EC 2.4.2.19 [nicotinate-nucleotide diphosphorylase (carboxylating)] the third step. In Escherichia coli, two of the residues that are involved in the [4Fe-4S] cluster binding appear to undergo reversible disulfide-bond formation that regulates the activity of the enzyme.
- EC 2.7.7.1, 18 Cofacteur : Néant
ATP + Nicotinate D-ribonucleotide <=> PP + Deamino-NAD+ (PPP primitif)
- EC 6.3.1.5 Cofacteur : Néant
ATP + Deamino-NAD+ + NH3 => AMP + PP + NAD+ (PPP primitif)
- EC 2.7.1.23 Cofacteur : Néant
ATP + NAD+ => ADP + NADP+ (PP primitif)
-
 Tetrahydrobioptérine
Tetrahydrobioptérine -
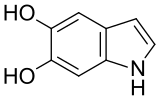 5,6-dihydroxyindole
5,6-dihydroxyindole -
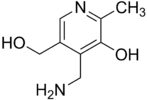 Pyridoxamine
Pyridoxamine -
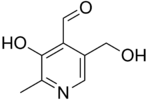 Pyridoxal
Pyridoxal
-Tetrahydrobiopterin_structure.svg)



Pyridoxal-P
[modifier | modifier le wikicode]- EC 2.6.1.30 Cofacteur: néant Pyridoxamine (serait hydrophobe) Métabolisme de B6
pyridoxamine + pyruvate <=> pyridoxal + L-alanine Pyridoxal (serait hydrophobe)
(EC 2.6.1.31 Cofacteur: néant
pyridoxamine + oxaloacetate <=> pyridoxal + L-aspartate)
- EC 2.7.1.35 Cofacteur: néant
ATP + Pyridoxal => ADP + Pyridoxal 5'_P (PP primitif)
- biosynthèse du Pyridoxal_P:
Voie des pentoses phosphates : 4.1.2.13(Zn) 3.1.3.11 2.2.1.1(B1)
Métabolisme de B6 : 1.2.1.72(NAD) 1.1.1.290(NAD) 2.6.1.52(Glu B6) 1.1.1.262(NAD)
voie des terpénoïdes : 2.2.1.7(D-Glyceraldehyde 3_P Pyruvate B1)
Métabolisme de B6: 2.6.99.2 1.4.3.5(O2 FMN) = Pyridoxal_P
- Récapitulatif: Schématiquement nous produisons à l'intérieur 3 produits fondamentaux à partir de 4 petites molécules dont 2 internes initiales et 2 externes et de 3 cofacteurs dont 2 sont externes et hydrophobes et le 3 ème petit, initial et interne.
| Réaction globale | Serine + Indole + D-Glycéraldéhyde-3P + O2 => Tryptophane + PRPP + NAD+
|
| Cofacteurs | Pyridoxal pyruvate Tétrahydrobioptérine
|
PRPP voie B1 :
[modifier | modifier le wikicode]- D-Glyceraldehyde 3_P + Glycerone_P
- Voie des pentoses phosphates : 412.13(Zn) 313.11 2211(B1) 2212 2211(B1) 2761(dATP) donne PRPP, 5427 donne Ribose-1P
- Voie des purines : 2421(A) 271.74(P) 2743(PPP) 2746(dNTP) donne ATP
- D-Glyceraldehyde 3_P + Acetaldehyde
- Voie des pentoses phosphates : 4124 5427 donne dRibose-1P
- Voie des purines : 2421(A) 3135(P) 2743(PPP) 2746(dNTP) donne dATP
- B1 est un cathion: Métabolisme de B1
- EC 3.5.99.2 : Thiamine + H2O <=> 4-Amino-5-hydroxymethyl-2-methylpyrimidine + 5-(2-Hydroxyethyl)-4-methylthiazole + H+ -----> les 2 produits sont supposés hydrophobes.
- EC 2.7.1.50 : 4-methyl-5-(2-hydroxyethyl)thiazole + ATP => 4-methyl-5-(2-phosphonooxyethyl)thiazole + ADP
- EC 2.7.1.49 : 4-amino-5-hydroxymethyl-2-methylpyrimidine + ATP => 4-amino-5-phosphonooxymethyl-2-methylpyrimidine + ADP
- EC 2.7.4.7 : (4-amino-2-methylpyrimidin-5-yl)methyl phosphate + ATP => (4-amino-2-methylpyrimidin-5-yl)methyl diphosphate + ADP
- EC 2513 : 2-methyl-4-amino-5-hydroxymethylpyrimidine diphosphate + 4-methyl-5-(2-phosphono-oxyethyl)thiazole => PP + thiamine phosphate
-
 4-Amino-5-hydroxymethyl-2-methylpyrimidine
4-Amino-5-hydroxymethyl-2-methylpyrimidine -
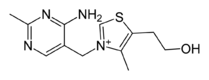 Thiamine
Thiamine -
 4-Methyl-5-(2-hydroxyethyl)-thiazole
4-Methyl-5-(2-hydroxyethyl)-thiazole


-thiazole.png)
Conclusion pour le choix de la voie vers PRPP et NAD+
[modifier | modifier le wikicode]La voie de l'Asp pour le quinolinate est très complexe : il faut de toutes les façons du FAD qui n'a pas d'origine hydrophobe ; ensuite Asp et fumarate viennent du cycle citrique qui est initialisé par l'oxaloacetate à partir du pyruvate qui vient du glyceraldehyde avec 4 réactions dont une de ferrodoxine mais pas de NAD ( 1.2.7.6 ) :
Contains tungsten-molybdopterin and iron-sulfur clusters. This enzyme is thought to function in place of glyceralde-3-phosphate dehydrogenase and possibly phosphoglycerate kinase in the novel Embden-Meyerhof-type glycolytic pathway found in Pyrococcus furiosus [1]. It is specific for glyceraldehyde-3-phosphate.
La 6.4.1.1, pyruvate vers oxaloacetate, est un verrou ( 4 cofacteurs : Acetyl-CoA, Manganese, Zinc, Biotine).
La voie de B1 pour PRPP avec le dihydroxyIndole pour le Quinolinate, reste la plus séduisante car elle utilise la biopterine équivalente de la tungsten-molybdopterine (1.2.7.6) et O2 (option de la 1.4.3.16).
- Pour démarrer la voie B1 il faut D-Glyceraldehyde 3_P et Glycerone_P.
Or à l'initialisation du métabolisme nous avons supposé dans l'hypothèse de la chimio-osmose prébiotique que le phosphate est libéré à l'intérieur ( ou utilisé directement à la surface interne de la membrane) par hydrolyse des liaisons esters :
3.1.1.4 puis 3.1.1.5 donnent ethanolamine_P_Glycerol et choline_P_Glycerol,
3.1.4.2 donne ethanolamine, choline et P_Glycerol,
3.1.3.21 donne le P et le glycerol.
3.1.3.27 libère le P,
3.1.4.3 (Zn++) directement P_glycerol,
P_ethanolamine et P_choline.

Pour le PRPP il faut considérer uniquement le glycérol. Mais celui-ci ne donne la glycerone qu'en présence de NAD(P)+ (1.1.1.6 et 156). L'hypothèse que je fais ici c’est que le glycérol et la glycérone diffuse facilement à travers la membrane ( à vérifier) et donc que l'initialisation de la production du NAD+ se fait par la glycérone puis tout le glycérol est transformé en glycérone grâce au NAD+.
La glycérone est ensuite piégée par le phosphate (2.7.1.29) puis isomérisée en D-Glyceraldehyde 3_P (5.3.1.1). qui ne demande pas de cofacteur ni de NAD+. On suppose qu’à l'initialisation ce sont P, PP et PPP qui sont utilisés.
- Acétaldéhyde : à partir de l'éthanol (diffuse facilement de l'extérieur, à vérifier)
1.1.1.1 NAD+ (Zn)
1.1.2.7 cytochrome cL :
A periplasmic quinoprotein alcohol dehydrogenase that only occurs in methylotrophic bacteria. It uses the novel specific cytochrome cL as acceptor. Acts on a wide range of primary alcohols, including ethanol, duodecanol, chloroethanol, cinnamyl alcohol, and also formaldehyde. Activity is stimulated by ammonia or methylamine. It is usually assayed with phenazine methosulphate. Like all other quinoprotein alcohol dehydrogenases it has an 8-bladed ?propeller? structure, a calcium ion bound to the PQQ in the active site and an unusual disulphide ring structure in close proximity to the PQQ. It differs from EC 1.1.2.8, alcohol dehydrogenase (cytochrome c), in having a high affinity for methanol and in having a second essential small subunit (no known function).
1.1.2.7 cytochrome c :
A periplasmic PQQ-containing quinoprotein. Occurs in Pseudomonas and Rhodopseudomonas. The enzyme from Pseudomonas aeruginosa uses a specific inducible cytochrome c550 as electron acceptor. Acts on a wide range of primary and secondary alcohols, but not methanol. It has a homodimeric structure [contrasting with the heterotetrameric structure of EC 1.1.2.7, methanol dehydrogenase (cytochrome c)]. It is routinely assayed with phenazine methosulphate as electron acceptor. Activity is stimulated by ammonia or amines. Like all other quinoprotein alcohol dehydrogenases it has an 8-bladed ?propeller? structure, a calcium ion bound to the PQQ in the active site and an unusual disulphide ring structure in close proximity to the PQQ.
- Finalement nous avons de l'extérieur :
O2 B1(2 parties) dihydroBioptérine dihydroxyindole glycérone éthanol
Initiateurs de l'initialisation du métabolisme
[modifier | modifier le wikicode]écrit le 28/07/11
- La thermodynamique du pétrole prébiotique devrait conduire à des sources d'énergie et des éléments sous forme de petites molécules dont l'énergie interne est la plus petite possible. Cependant les initiateurs fabriqués dans la poche de pétrole, grace notamment aux surfaces minérales, peuvent être complexes. La seule règle qu’ils doivent suivre c’est de rentrer dans le liposome. Pour cela il faut qu’ils soient hydrophobes.
Au contraire des méthanogènes qui utilisent comme source d'énergie des minéraux et du comme source de carbone, les méthanotrophes des poches de pétrole peuvent utiliser le méthane et les hydrates de carbones comme source d'énergie et de carbone à la fois: méthane, méthanol, éthanol, propanol... Ceci correspond au processus de FT qui produit d’abord des alcools primaires. De ce point de vue là le glycérol est la molécule minimale pour fabriquer les lipides.

- Les initiateurs(précurseurs) du métabolisme vont utiliser de petites molécules pour les aa:
- d’abord pour les aa hydrophobes, ils sont issus des amines(FT comme alcools): méthylamine, éthyl-, isopropyl-, butyl-, pentyl-, pour respectivement gly, ala(et beta-alanine), val, pro, ile et leu.
- issus de diamines: butyl-diamine pour arg et pentyl-diamine pour lys.
- issus d'une amine + alcool ou thiol: éthyl( ser, cys, éthanolamine, cystéaimne ), isopropyl( pro ), propyl( met ).
- Les précurseurs peuvent être plus complexes.
On sait que FT produit avec le temps des cycles aromatiques.
- les 4 aa aromatiques: leur précurseurs vont être sous forme d'alcool comme ceux de certains cofacteurs: histidinol(his), phénol(tyr qui est lui-même précurseur de phe), indole qui donnera trp en réaction avec ser.
- les bases an: ce sont les bases directement puisqu'elles sont hydrophobes.
- les cofacteurs: alcool( pantothénol pour CoA, pyridoxine pour B6, indole-diol pour NAD+, méthanol-thiazole et éthanol-pyrimidine pour B1, dihydroptéridine pour THF, tétrahydrobioptéridine pour BH4 lui-même), amine précurseur(?) de THMPT, acides longs donc plus hydrophobes( pimelate pour biotine et acide lipoique), les isoprènes pour pyroles et quinones(+ benzoateOH pour les quinones).
- les aa et les cofacteurs sans précurseurs sont synthétisés par le métabolisme TCA( asp, glu, asn, gln ) et celui des flavine (FAD FMN).
- L'origine des AN, des aa et de la membrane sont intimement liés dans les cofacteurs: CMP-membrane, PRPP donne AN, His et NAD. Ensuite on a les liaisons H entre aa et AN, hydrophobie entre ionophores, cofacteurs et membrane.
- Les cytochromes avec Fe++ ou bien Fe++, Zn++ et Cu++ seuls semblent être les plus primitifs:
- Methanol(ethanol) donne formaldehyde(acetaldehyde: EC 1.1.2.7 et 8 (cytochrome cL ou c, quinone et PQQ, localisation périplasmique).
- Méthane donne méthanol: EC 1.14.18.3 (quinol et Cu++, localisation membranaire).
- BH4 et indole-diol donnent quinolinate: EC 1.13.11.23 (O2), EC 1.14.16.3 (O2, BH4, Fe++), EC 1.13.11.6 (O2, Fe++); localisation membranaire?
- imino-aspartate et GOP donnent quinolate: EC 2.5.1.72 (cluster 4Fe-4S, localisation plastide chez les eucaryotes, mais E.Coli?); la réaction avant celle-ci réduit l'asp en imino-aspartate, EC 1.4.1.21(NAD+) ou EC 1.4.3.16(FAD) et à la place d'asp?
- GAP et GOP donnent fructose-PP: EC 4.1.2.13 (Zn++, localisation cytoplasme chez les eucaryotes, mais E.Coli?).
Diffusion à travers la membrane
[modifier | modifier le wikicode]écrit le 03/08/11
-poser le problème du transport de part en part ou fixation dans la couche d'huile.
-les aa ester-méthylés ont un coefficient de perméabilité x par 100.
-problème de la perméation de l'eau.
-nombre de molécules par liposome: lipd bilayer-NCBI.
-le cholesterol a une tête OH.
-les ah seraient plus hydrophobes que les aa.
- Deamer (1992)[3] :
EPC : egg phosphatidylcholine, (wiki: phosphatidylcholine), palmitic saturé(16C) ou margaric saturé(17) et oleic(18) insaturé au 9Z.
DMC : dimyristoylPC (14) saturé.
Tableau coefficient de perméabilité P 10-13 cm/s.
| aa | EPC | DMC | propriété du radical |
|---|---|---|---|
| Gly | 57 | 200 | neutre |
| Ser | 55 | 160 | polaire |
| Lys | 51 | 190 | chargé |
| Trp | 4100 | - | aromatique |
| Phe | 25 | - | hydrophobe |
| Phosphate | 5 | - | anion |
| Na+ K+ | 5 | - | cation |
- Le pH a peu d'influence sur le coefficient P des aa testés.
- Le coefficient P est inversement proportionnel à la longueur de la chaine aliphatique.
- Le phosphate a un coefficient P entre 10-13 et 10-12 cm/s.
- Les aa neutres, polaires et chargés ont un coefficient P proche des cations Na+ et K+.
- (note: pourquoi Phe a un P comme ces derniers alors que les hydrophobes passent plus facilement; s'attarde-t-il dans la couche d'huile? Et y resterait comme les aliphatiques 100% hydrophobes?).
- Cette observation(article) suggère que la perméabilité des aa neutres, polaires et chargés est due aux fluctuations et aux défauts transitoires de la bicouche, plus qu'au coefficient de partage, octanol/eau, ou bien la barrière énergétique de Born (voir article sur les aa aromatiques).
- Chakrabarti (1994) review[4].
- La perméabilité croit avec la réduction de la chaine aliphatique(5 fois).
- La perméabilité croit avec la charge surfacique pour les aa de charge opposée: choline/anion.
- La perméabilité croit avec la teneur de cholestérol de la membrane.
- Les aa neutres, polaires et chargés ont un coefficient P proche des cations Na+ et K+.
- Les aa hydrophobes sont 100 fois plus perméables que les aa hydrophiliques, reflétant les valeurs élevées de leurs coefficients de partage, octanol/eau( Phe!!! voir ma note pour Dreamer 1992).
- La perméabilité des aa ester-méthylés (comme Lys) est très élevée et est réduite drastiquement par le pH à cause du gradient pH qui s'établit. Il a été établit que l'ester de Lys-methyl et des peptides courts modifiés ont un un coefficient P d'environ 10-2 cm/s comme les molécules neutres (déprotonées).
- La distribution des charges (sur la surface du liposome?) altère drastiquement la perméation des dipeptides modifiés.
- Ceci serait lié au transport membranaire primitif!!!
- Deamer (1994)[5]:
- Les transports des aa et autres nutriments, comme le phosphate, sont 105 plus lents que les transports facilités.
- Soit les membranes primitives sont intrinsèquement plus perméables (longueur de chaine) ou bien un mécanisme quelconque facilite le transport.
- Le transport d'espèces chimiques neutres est un de ces mécanismes (moi je propose les précurseurs du métabolisme, non chargés et stables).
- Les aa transformés par méthylation ont un coefficient P 1010 supérieur à celui des espèces chargées.
- Cette perméation associée au gradient pH transmembranaire doit accroitre significativement le taux de transport unidirectionnel, direction définie par le gradient.
- Ces gradients de pH ont dû exister chez les vésicules primitives.
- Ces mécanismes de transport(sans protéines) doivent jouer encore un rôle in vivo pour la translocation de certaines protéines(peptides courts): séquences de signalisation, toxines(mes ionophores) et des protéines thylakoïdales(à vérifier).
- En introduction de cet article: il a été calculé que le transport des aa pour de simples organismes comme E.Coli est de 10-2 nmol/s.cm2
en assumant que le poids moyen d'un aa est de 150 et qu’il faut 20 min pour que la bactérie double son contenu en protéines. Le même calcul pour un liposome donne un flux approximatif de 10-7 nmol/s.cm2.
- Deamer (1983)[6]:transport des H+. Le gradient pH diminue la vitesse de transport des protons hydroxydes (H3O+). Pour une différence de pH de 3 unités la vitesse est de 10-9 cm/s et pour un faible gradient elle est de 10-4 cm/s.
- Cours Jakubowski (2010):
- Tableau 1. Quelques valeurs du coefficient P de perméabilité pour des vésicules synthétiques adaptées de la figure "Permeability coefficients".
- Tableau 2. Comparaison entre vésicule synthétique et erythrocyte humain intact pour illustrer entre transport actif du glucose et passif du mannitol: valeurs expérimentales en cm/s.
- Le cours cite la valinomycine comme ionophores qui sont des transporteurs non protéiques de cations dans le sens décroissant du gradient de concentration.
Tableau 1. Coefficient de perméabilité P en cm/s.
| Na+ | 10-12 | glucose | 5.10-8 | indole | 5.10-4 |
| K+ | 5.10-11 | Trp | 10-7 | éthanol, eau, acide acétique |
entre 10-3 et 10-2 |
| Cl- | 10-10 | glycérol, urée | 5.10-6 | acide hexnoïque | 10-0 |
Tableau 2. Transport actif/passif en cm/s:
| molécule | D-glucose | D-mannitol |
|---|---|---|
| Vésicule synthétique | 2.4x10-10 | 4.4x10-11 |
| Diffusion calculée | 4x10-9 | 3x10-9 |
| érythrocyte humain | 2.0x10-4 | 5x10-9 |
- Article Lipid bilayer sur wikipedia:
- Flopase: pgr assymétrie
- H2O, anions: pgr passive diffusion
- reproduction des liposomes: pgr containment and separation
- Les vésicules synthétiques: pgr model systems
- Archées à membrane simple couche: pgr containment and separation
Références
[modifier | modifier le wikicode]- ↑ Sandra Pizzarello & Arthur L. Weber: Stereoselective Syntheses of Pentose Sugars Under Realistic Prebiotic Conditions. Orig Life Evol Biosph (2010) 40:3–10 DOI 10.1007/s11084-009-9178-1
- ↑ Hideaki Tsukahara, Ei-ichi Imai, Hajime Honda, Kuniyuki Hatori and Koichiro Matsuno: Prebiotic oligomerization on or inside lipid vesicles in hydrothermal environments. Origins of Life and Evolution of the Biosphere 32: 13–21, 2002
- ↑ Ajoy C. Chakrabartia and David W. Deamer: Permeability of lipid bilayers to amino acids and phosphate. Biochimica et Biophysica Acta (BBA) - Biomembranes Volume 1111, Issue 2, 9 November 1992, Pages 171-177 ScienceDirect
- ↑ A. C. Chakrabarti: Permeability of membranes to amino acids and modified amino acids: Mechanisms involved in translocation. Biomedical and Life Sciences; Amino Acids Volume 6, Number 3, 213-229, DOI: 10.1007/BF00813743. ScienceDirect
- ↑ Ajoy C. Chakrabarti and David W. Deamer: Permeation of membranes by the neutral form of amino acids and peptides: Relevance to the origin of peptide translocation; Biomedical and Life Sciences Journal of Molecular Evolution Volume 39, Number 1, 1-5, DOI: 10.1007/BF00178243 ScienceDirect
- ↑ David W. Deamer and J. Wylie Nichols: Proton-hydroxide permeability of liposomes; Proc. Natl Acad. Sci. USA Vol. 80, pp. 165-168, January 1983 Biophysics